3.2. Система “цепочка
артериальных сосудов”
3.2.1. Представление системы
Согласно
используемому нами алгоритму построения систем [136, 137], система “цепочка
артериальных сосудов” включает в себя в качестве элементов цепочку последовательных
сосудов, по которой движется отдельный эритроцит, кровь в каждом из них и давление, приложенное к этим
сосудам. Отношением единства этих элементов являются функциональная зависимость
между ними. Законы композиции системы будут представлены нами ниже.
3.2.2. Условия оптимального
движения крови в сосуде
В любом сосуде
расход энергии, связанный с движущейся кровью, разделяется на следующие составляющие
[97]:
1) расход
энергии, теряемой в результате вязкой диссипации;
2) расход
энергии, потребляемой в связи с генерацией клеточных элементов крови взамен погибающих;
3) расход
энергии, затрачиваемой особью на передвижение в связи с необходимостью
“переносить” кровь;
4) расход
энергии на метаболизм клеточных элементов крови.
Затраты
энергии, представленные в 2)-4), составляют “стоимость” крови. С.Мюррей [239]
отмечает что четвертый фактор “стоимости” существенно меньше второго и третьего. В дальнейшем будем обозначать энергию,
расходуемую на перфузию сосуда, “перфузионной”, а энергию, связанную со “стоимостью” крови, - “химической”.
При заданном
кровотоке в сосуде с увеличением диаметра расход “перфузионной” энергии падает, а
“химической”, наоборот, возрастает; следовательно, для некоторого диаметра суммарный расход энергии в сосуде
будет минимальным. В соответствии с принципом “минимальности работы”, выдвинутым
Мюрреем [239], заданный кровоток в сосуде q связан с его диаметром d и параметрами крови
оптимальным соотношением
q/d3=
(p/32)![]() ,
,
где x - вязкость крови, b - расход “химической”
энергии в единичном объеме крови за единицу времени.
C другой
стороны, легко показать, что
q/d3=
(p/4)l/dt
= a,
где d, l, t - соответственно
диаметр и длина сосуда, время пребывания эритроцитов в сосуде, a - коэффициент оптимальной перфузии. Очевидно, что
минимальность суммарного расхода энергии в коронарном сосуде существует лишь в том
случае, если
l/dt
= ![]() /8.
(3.1)
/8.
(3.1)
В левой части
этого равенства представлены параметры сосуда, в правой - параметры крови. Элементом,
функционально объединяющим обе части равенства, является диаметр сосуда, поскольку параметры крови x и b зависят от величины d
[152, 153].
3.2.3. Анализ
архитектонических и гемодинамических параметров в золотом режиме гипертензии
Система
докапиллярного транспорта кислорода представляет собой цепочку сосудов, по
которой отдельный эритроцит движется от аорты до капилляра. Цепочка состоит из
последовательности сосудистых тройников (см. рис. 3.1). Длина тройника, или звена цепочки, равна расстоянию по
стволу от вершины угла разветвления при котором образовался этот ствол, до
вершины угла, при котором он распался на ветви (рис. 3.2) Конструкционные особенности тройника в золотом режиме гипертензии
можно представить тремя коэффициентами [166]:
1) коэффициент ветвления
k(bзс ) = [S1(bзс) + S2(bзс)]/SС(bзс),
(3.2)
2) коэффициенты
относительного сечения ветвей
H1(bзс) = S1(bзс)/[S1(bзс) + S2(bзс)], (3.3)
H2(bзс) = S2(bзс )/[S1(bзс) + S2(bзс)], (3.4)
где SС(bзс), S1(bзс), S2(bзс), - площади поперечных
сечений ствола, 1 и 2 ветвей в золотом режиме гипертензии. В выражениях (3.2) - (3.4) представлены
средние за сердечный цикл (средние) значения площадей.
|
|
Рис. 3.1. Цепочка сердечных сосудов от аорты до капилляра, по которой
движется отдельный эритроцит.
Представленная
в книге К.А. Шошенко и др. [166] экспериментальная зависимость k=f(H) позволила
установить, что для артериального участка коронарного русла k=1,10-1,20. В этой же работе авторами
теоретически доказано, что величинам 1<k£1,26 в тройниках соответствует
оптимальный режим “минимальной работы”, установленный Мюрреем [238, 239]. Очевидно,
что движение крови по артериальному коронарному руслу имеет оптимальную основу.
|
|
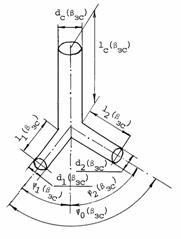
Рис. 3.2. Схема сосудистого тройника. dс(bзс), d1(bзс), d2(bзс), lс(bзс), l1(bзс), l2(bзс) - соответственно
диаметр и длина ствола и 1 и 2 ветвей, j1(bзс), j2(bзс) - углы отклонения 1 и 2 ветвей, j0(bзс) - угол разветвления в золотом режиме гипертензии.
Рассмотрим
соотношения диаметров и длин ствола и одной из ветвей тройника (последовательных
звеньев цепочки) с привлечением коэффициентов k(bзс ) и H(bзс ).
Для оценки
взаимосвязи между длиной и диаметром сосуда введем конструктивный параметр с(bзс )=l(bзс )/d(bзс ). К.А.Шошенко и др. [166]
отмечает, что распределение этого параметра по цепочке коронарных сосудов аналогично и для
млекопитающих, и для рыб, и для земноводных. Установлены особенности распределения
величины конструктивного параметра при движении по цепочке. Так для участка цепочки, где d(bзс)>50 мкм, c(bзс)=3, а для участка в
диапазоне d(bзс)<50
мкм - c(bзс)=4-6. Всякий ствол и одна
из его ветвей являются последовательными i- и i+1-звеньями цепочки сосудов, по которой движется отдельный
эритроцит. Для любых i- и i+1-звеньев цепочки имеют место отношения [152]
d i+1(bзс) =![]() di(bзс), (3.5)
di(bзс), (3.5)
l i+1(bзс) = ![]() [ci+1(bзс)/c i(bзс)]l i(bзс), (3.6)
[ci+1(bзс)/c i(bзс)]l i(bзс), (3.6)
где ki+1(bзс) - коэффициент ветвления
тройника при переходе от i- к i+1-сосуду, Hi+1(b ) - относительное сечение i+1-сосуда, ci(bзс), c i+1(bзс) - коэффициенты конструкции
i- и i+1-сосудов в золотом режиме гипертензии.
Покажем, что
для всех сосудов цепочки величина коэффициента оптимальной перфузии a(bзс) неизменна. С этой целью
рассмотрим изменения параметров, связанных с a(bзс). C учетом (3.5-3.6) в соседних сосудах
Vi+1(bзс)=(p/4)(di+1(bзс))2li+1(bзс)=
=(ki+1(bзс)Hi+1(bзс))3/2[ci+1(bзс)/ci(bзс)]Vi(bзс), 3.7)
где Vi(bзс), Vi+1(bзс) - объемы i- и i+1-сосудов
в золотом режиме гипертензии. В режиме “минимальной работы” кровотоки в стволе и ветви связаны выражением
[166]
q i+1(bзс) = (k i+1(bзс)H i+1(bзс))3/2 qi(bзс), (3.8)
где qi(bзс), q i+1(bзс) - кровотоки в i- и
i+1-сосудах в золотом режиме.
Исходя из
(3.8) и (3.9),
t
i+1(bзс)
= V i+1(bзс)/q i+1(bзс) = [c i+1(bзс)/ci(bзс)]ti(bзс), (3.9)
где ti(bзс), ti+1(bзс) - время пребывания
эритроцитов в i- и i+1 -сосудах в золотом режиме гипертензии.. Для некоторого i-сосуда
ai(bзс) = (p/4)li(bзс)/di(bзс)ti(bзс),
где ai(bзс) - коэффициент оптимальной
перфузии i-сосуда. Соответственно, для i+1-сосуда с учетом (3.5), (3.6) и (3.9)
a i+1(bзс) = (p/4)l
i+1(bзс)/d i+1(bзс)t i+1(bзс) =
= (p/4)![]() [c i+1(bзс)/ci(b )] li(bзс)/
[c i+1(bзс)/ci(b )] li(bзс)/
/![]() di(bзс)´
di(bзс)´
´![]() [ci+1(bзс)/ci(bзс)]ti(bзс) = ai(bзс). (3.10)
[ci+1(bзс)/ci(bзс)]ti(bзс) = ai(bзс). (3.10)
Таким образом,
несмотря на наличие “нестандартных” коэффициентов конструкции в терминальных
микрососудах (c>3) коэффициент перфузии постоянен всей длине цепочки [152],
a(bзс)
= 109,5 с-1.
Покажем, что
давление, приложенное к каждому сосуду цепочки, является постоянным для всех сосудов
цепочки.
Кажущаяся
вязкость крови в соответствии с феноменом Фареуса-Линдквиста зависит от диаметра
сосудов. При сопоставлении экспериментальных зависимостей x=f(d) [188] и с=j(d) [166] нами получено обратное
соотношение этих параметров для соседних сосудов [152, 153],
x i+1(bзс)/xi(bзс) @ сi(bзс)/с i+1(bзс), (3.11)
где xi(bзс), x i+1(bзс) - кажущаяся вязкость в i-
и i+1-сосудах в золотом режиме гипертензии.
Давление, приложенное к
i-сосуду,
DPi(bзс ) = qi(bзс)/Gi(bзс),
где DPi(bзс ), Gi(bзс)- соответственно давление,
приложенное к i-сосуду, и проводимость i-сосуда в золотом режиме. С учетом (3.6) и (3.11) проводимость
i+1-сосуда
Gi+1(bзс), = pd i+1(bзс)4/128x i+1(bзс)l i+1(bзс) =
= p(k i+1(bзс)H i+1(bзс))2(d (bзс))4/128(ci(bзс)/
/c i+1(bзс))xi(bзс)(c i+1(bзс)/ci(bзс))´
´![]() li(bзс ) =
li(bзс ) =
= [k i+1(bзс)H i+1(bзс)]3/2Gi(bзс). (3.12)
Поскольку кровоток и
проводимость в i+1-сосуде по отношению к i-сосуду изменяются в (kH)3/2 раз,
то очевидно, что
DPi+1(bзс) = DPi(bзс). (3.13)
Для любого артериального сосуда сердца [152]
DP(bзс) = 2,85 мм рт. ст.
Рассмотрим
энергетическую основу транспорта крови по цепочке артериальных сосудов. Во
всяком i-сосуде за единицу времени общий расход энергии складывается из потерь на
перфузию и “накладных расходов”. Последние обусловлены расходом мощности на воспроизводство “изношенных”
эритроцитов и расходом мощности, связанным с весом и переносом крови [97].
Поскольку “накладные” потери в большей степени связаны с эритропоэзом, то будем в дальнейшем энергию, соответствующую
этим потерям, обозначать “химической”. Затраты мощности во всяком i-сосуде равны сумме затрат “перфузионной” и
“химической” мощностей.
Механическая,
“перфузионная”, мощность в i-сосуде
Wпi(bзс) = qi(bзс)2128xi(bзс)li(b )/pdi(bзс)4, (3.14)
где Wпi(bзс) - перфузионная мощность в
i-сосуде в золотом режиме. Легко показать с учетом (3.5), (3.6), (3.8) и (3.11), что для i+1-сосуда
Wп(i+1)(bзс) = (k i+1(bзс)H i+1(bзс))3/2 Wпi(bзс). (3.15)
“Химическая” мощность в i-сосуде
Wxi(bзс) = (p/4)bi(bзс)di(bзс)2li(bзс), (3.16)
где bi(bзс)- скорость потребления
“химической” энергии в единичном объеме крови в золотом режиме. Исходя из (3.1)
bi(bзс)= 64xi(bзс)li(bзс)2/di(bзс)2ti(bзс)2. (3.17)
Поскольку независимо от
калибра сосуда l(bзс)/d(bзс)t(bзс)=const, то очевидно, что величины x и b во всяком сосуде
изменяются в равной степени. С учетом этого обстоятельства легко показать, что
Wx(i+1)(bзс) = (k i+1(bзс)H i+1(bзс))3/2Wxi(bзс). (3.18)
Из сравнения выражений (3.15) и (3.18) следует
вывод, что
Wп(i+1)(bзс) / Wx(i+1)(bзс) = Wпi(bзс)/Wxi(bзс).
Очевидно, что для всех
артериальных сосудов сердца существует постоянная безразмерная величина
g(bзс) = Wп(bзс)/Wx(bзс) = const.
Таким образом,
в золотом режиме гипертензии по всей цепочке артериальных сосудов для каждого звена
независимо от изменений d, l, t, x и b сохраняются неизменными три постоянных величины (инварианта): DP(bзс), a(bзс) и g(bзс). Законы симметрийных преобразований, при которых
эти инварианты сохраняются, могут быть получены из законов изменения d, l, t, x и b. Безразмерное отношение g(bзс) является инвариантом, как
будет показано нами в дальнейшем, не только для артерий, но и для капилляров, и не только для покоя, но и при
любом уровне гипертензии.
3.2.4. Анализ
кислородных параметров в золотом режиме гипертензии
Изменение
кислородных параметров по цепочке связаны, как мы увидим дальше, с диаметром каждого
сосуда и временем пребывания эритроцита в сосуде. Несомненно, что наибольший интерес представляют терминальные
звенья цепочки, поскольку здесь в основном происходит докапиллярная диффузия
кислорода в ткани. Произведем вычисление величин диаметров, начиная от капилляра по направлению к аорте; примем
диаметр капилляра d(bзс)=5,5 мкм [181]. Величину диаметра соседнего сосуда
определяем по формуле [152],
dm+1(bзс) = dm
(bзс)/![]() ,..
,..
где dm(bзс), dm+1(bзс) - диаметры соседних
сосудов цепочки по направлению от капилляра к аорте в золотом режиме гипертензии. Расчетные значения диаметров
первых 10 сосудов приведены в табл. 1 (столбец 2) [152].
Время
пребывания эритроцита на артериальном участке сосудистого тракта составляет
(15-20)% от общего времени [126]. В покое общее время пребывания эритроцита в
коронарном русле собак при нормальном среднем давлении в аорте, равном 98 мм рт. ст., составляет 4 с [184].
Очевидно (см. (3.9)), что для соседних звеньев цепочки с одинаковым значением с(bзс)=3 время пребывания постоянно. Выражение (3.9)
справедливо для любых двух, не обязательно соседних, звеньев цепочки. Следовательно, приняв в качестве исходного диаметр
некоторого сосуда, у которого с0(bзс)=3, а время пребывания в нем за исходную величину
для расчетов,
мы могли бы рассчитать по отношению к этому сосуду относительные изменения
пребывания во всех остальных сосудах от капилляра до аорты. По произведенным нами
расчетам число звеньев в артериальной цепочке собак n=28 [152]. Полученные нами значения времен пребывания в первых
10 звеньях цепочки, начиная от капилляра, представлены в столбце 4 таблицы 1. Во всех артериальных сосудах, за
исключением первых 7 звеньев, t(bзс)=21,5
мсек.
Таблица
1
Величины
гемодинамических и кислородных параметров в терминальных артериолах
|
m |
dm(bзс),мкм |
cm(bзс) |
tm(bзс), c |
Um(bзс), %/c |
DHbO2m(bзс), % |
|
1. |
7,2 |
6 |
0,043 |
127,6 |
5,49 |
|
2. |
9,3 |
6 |
0,043 |
74,2 |
3,18 |
|
3. |
12,2 |
5 |
0,0359 |
42,9 |
1,54 |
|
4. |
15,9 |
5 |
0,0359 |
24,9 |
0,89 |
|
5. |
21,4 |
4 |
0,0286 |
13,7 |
0,39 |
|
6. |
28,9 |
4 |
0,0286 |
7,5 |
0,21 |
|
7. |
37,6 |
4 |
0,0286 |
4,1 |
0,12 |
|
8. |
49,2 |
3 |
0,0215 |
2,3 |
0,05 |
|
9. |
64,2 |
3 |
0,0215 |
1,2 |
0,02 |
|
10. |
83,7 |
3 |
0,0215 |
0,7 |
0,01 |
Примечания: m
- номер артериального сосуда цепочки, начиная от капилляра; tm(bзс) - время пребывания
эритроцита, Um(bзс) - скорость отдачи кислорода отдельным эритроцитом, DHbO2m(bзс) - количество кислорода,
отданного отдельным эритроцитом за время пребывания в m-сосуде в золотом режиме
гипертензии.
Теперь
перейдем к непосредственному расчету кислородных параметров. Эритроциты
отдают кислород не только в капиллярах, но и мелких артериях [62, 196] и венулах [193].
Установлено [196], что существует практически линейная зависимость между толщиной стенки микрососуда и
разностью DрО2 на его
внешней и внутренней поверхностях. Из анализа экспериментальных данных [196, 220] следует, что отношение
толщины стенки сосуда к его диаметру - величина, приблизительно одинаковая для всех
микрососудов. Как следствие этого, отношение площадей, занятых кровью и сосудистым
материалом в поперечном сечении всякого сосуда, имеет одно и то же значение. Очевидно, что одинаковое
отношение между изменениями толщины стенки и величины DрО2 возможно лишь тогда, когда
каждое сечение любого микрососуда отдает за единицу времени одно и то же количество кислорода.
Очевидно, что вклад отдельного эритроцита в общую диффузию кислорода из поперечного сечения
будет тем больше, чем меньше площадь последнего. Иными словами, при переходе
эритроцитов из одного сосуда в другой
скорость отдачи кислорода отдельным эритроцитом изменяется обратно пропорционально
изменению площади поперечного сечения. В капилляре эритроцит, естественно, отдает кислород с максимальной скоростью.
Исходя из этих рассуждений,
Um(bзс) =Uk (bзс)Sk(bзс)/Sm(bзс), (3.19)
где Um(bзс), Uk(bзс), Sm(bзс), Sk(bзс) - соответственно скорость отдачи кислорода отдельным
эритроцитом и площадь поперечного сечения в m-артерии и капилляре в золотом
режиме гипертензии.
По расчетам
[61] напряжение кислорода на артериальном конце капилляра в тканях
составляет 50-60 мм рт. ст. Примем для дальнейших расчетов в исходном режиме рО2(bзс)=50 мм рт. ст., а в первой коронарной артерии рО2=95
мм рт. ст. По кривой диссоциации, взятой нами из книги [129] (рис. 3.3), этому интервалу соответствует
разность насыщения кислородом эритроцита DHbO2=12%. При прохождении m-сосуда
разность насыщения кислородом эритроцита кислородом в этом сосуде
DHbO2(bзс) = Um(bзс)tm(bзс) = Uk(bзс)Sk(bзс)tm(bзс)/Sm(bзс).
Соответственно, для всего артериального участка
DHbO2(bзс) = Uk(bзс)Sk(bзс) ![]() ( bзс)/Sm(bзс). (3.20)
( bзс)/Sm(bзс). (3.20)
|
|
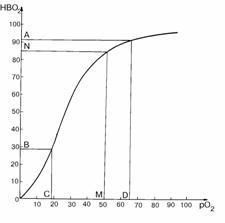
Рис. 3.3. Кривая насыщения гемоглобина человека [126]. По оси абсцисс -
pO2 в мм рт. ст.; по оси ординат - HbO2 - в %. Точки A,
B, C, D - границы логарифмического участка кривой (см. рис. 4.3). Интервал CD -
pO2 = 19-65 мм рт. ст.; интервал AB - HbO2 =28,7-91,8%;
точкам M и N соответствуют pO2 = 50 мм рт. ст. и HbO2
=85% на артериальном конце открытого сердечного капилляра в золотом режиме гипертензии.
Скорость
отдачи кислорода эритроцитом в капилляре в исходном режиме можно определить по
формуле
Uk(bзс) = (DHbO2(bзс)/ Sk(bзс))/ ![]() ( bзс)/Sm(bзс).
( bзс)/Sm(bзс).
Для вычисления Uk(bзс) вполне достаточно первых
10 звеньев цепочки, начиная от капилляра [152]. Приняв DHbO2(bзс)=12%, Sk(bзс)=23,7*10-8 см2
и используя значения dm(bзс) и tm(bзс), приведенные в табл. 1, можно
вычислить величины Uk(bзс), Um(bзс) и DHbO2m(bзс); последние приведены в
табл. 1 (см. столбцы 5 и 6). Расчетное значение Uk(bзс)=215%/с практически равно
максимальной скорости отдачи - 220%./c. Эта величина была получена при определении скорости
диссоциации оксигемоглобина для движущихся в плазме эритроцитов с имитацией потребления
кислорода [162, 243]. Экспериментально установлено [260], что скорость отдачи
кислорода из сосуда практически не меняется при значительных изменении
кровотока и гематокрита.
Интересно
отметить, что последовательность значений Um(bзс) (см. табл. 1), в том числе и
Uk(bзс),
по мере приближения к капилляру все более приобретает форму ряда Фибоначчи: 2,49*10; 4,29*10; 7,42*10; 12,76*10;
21,5*10 (для сравнения числа классического ряда Фибоначчи: 3, 5, 8, 13, 21). Эта последовательность указывает
на то, что изменение отдачи кислорода единичным эритроцитом в терминальных артериолах и капилляре происходит по
аналогии с рядом Фибоначчи. Поскольку размеры терминальных микрососудов у различных
видов млекопитающих имеют поразительное сходство [64], то очевидно, что скорость отдачи кислорода отдельным
эритроцитом в данном m-микрососуде одинакова для всех животных независимо от их
размера. Следовательно, скорости “разряда” эритроцитов в терминальных артериолах являются
инвариантными величинами.
Любопытно
отметить в связи с этим соотношение между парциальным давлением кислорода во вдыхаемом
воздухе рО2 =156 мм рт. ст. [13] и напряжением кислорода в артериальной крови рО2=95 мм рт. ст.; падение рО2
в легких составляет 61 мм рт. ст. Очевидно, что величины рО2 , соответствующие
свободной и химически связанной форме кислорода в организме, приблизительно соотносятся между собою по золотой
пропорции - 156:95 = 95:61@ 1,6.
3.2.5.
Особенности организации прекапиллярного участка в золотом режиме гипертензии
Из анализа
столбцов табл. 1 очевидно, что время пребывания и скорость отдачи кислорода
отдельным эритроцитом возрастают по мере его приближения к капилляру. Известно
[176], что для терминальных артериол сердца характерно постоянство или незначительное изменение
размеров в течение всего кардиоцикла. В первых от капилляра 7 сосудах эритроцит
отдает более 98% из общего количества кислорода, отданного на всем артериальном участке (см. табл. 1).
Дальше десятого звена цепочки количеством кислорода, отдаваемого отдельным эритроцитом,
при расчетах можно пренебречь.
Отличительной
особенностью пассажа эритроцитов по цепочке является увеличение времени пребывания по
мере приближения к капилляру (см. табл. 1). Этот феномен имеет большой физиологический смысл. Если
бы время пребывания было неизменным и всюду равным 0,0215 с, то молекулы
оксигемоглобина до капилляра не успели бы “разрядиться” до величины рО2(bзс)=50 мм рт. ст. [152]. Благодаря
“рациональной” временной организации прохождения эритроцитов в терминальных
артериолах молекулы оксигемоглобина при поступлении в капилляр имеют оптимальное
значение рО2(bзс)=50 мм рт. ст. Следует обратить внимание на то, что прекапиллярные ветвления
весьма близки к полной симметрии [46]. Вследствие этого эритроциты в ветвях терминальных
тройников отдают равное количество кислорода и входят в капилляры с приблизительно равными рО2. В
противном случае, т.е. при значительной асимметрии терминальных бифуркаций, степень
““разряда” эритроцитов в ветвях была бы различной. Поскольку “разряд” соответствует медленно спадающему
участку кривой насыщения гемоглобина (рис.3.3), то эритроциты входили бы в капилляры со значительными отклонениями
от рО2=50 мм рт. ст. Аналогичность величин рО2 на
артериальном и венозном концах для всех капилляров, как будет показано нами в 4
главе, имеет большое физиологическое значение, поскольку позволяет обеспечить одинаковые условия снабжения
кислородом по всей длине мышечных волокон.
Таким образом,
прекапиллярные сосуды являются своего рода переходной конструкцией, где происходит
предварительная организация оптимальных условий диффузии кислорода в капилляре. Расположение “ряда”
скоростей диффузии кислорода в прекапиллярных звеньях цепочки происходит в
соответствии с рядом чисел Фибоначчи. Этот “ряд” скоростей является инвариантом для всех млекопитающих независимо от
их размера.
3.2.6. Законы
композиции системы в режимах гипертензии
В общем виде
закон структурно-динамической композиции любого i-звена в некотором режиме гипертензии
можно представить выражением
DPi(b)ti(b) = 32xi(b)[li(b)/di(b)]2 . (3.21)
В этом выражении левая часть
отображает динамические параметры, т.е. давление, приложенное к i-сосуду, и
время пребывания эритроцитов в i-сосуде. В правой части представлены структурные параметры - кажущаяся
вязкость крови, длина и диаметр i-сосуда.
В золотом
режиме гипертензии для “стандартных” сосудов имеем li(bзс)/d1(bзс)=3 [166], xi(bзс)=3*10-2г см-1с-1 [212] и расчетные величины DPi(bзс)=2,85 мм рт. ст.=380 г см-2с-2
и ti(bзс)=21,5*10-3c.
Подставив эти данные, полученные экспериментальным путем и в результате
расчетов, в (3.21), получим незначительное расхождение между левой и правой частями этого равенства, что
указывает на достаточно точное определение расчетных величин DPi(bзс) и ti(bзс). В режимах гипертензии параметры DPi(b), ti(b), xi(b), di(b) и li(b) изменяются в соответствии с законами их
преобразования. Эти законы будут представлены нами при последующем анализе
гемодинамических параметров.
Рассмотрим
изменения структурных параметров цепочки сосудов при возрастании среднего аортального
давления в b раз. Предпринятый нами анализ относительных изменений диаметров и объемов аорты и крупных
коронарных артерий показал, что при гипертензии [151 152]
di(b) = b1/3di(bзс), (3.22)
li(b) = b1/3 li(bзс ), (3.23)
где di(b), li(b) - соответственно диаметр и
длина i-сосуда в заданном режимах гипертензии. Следует отметить, что в (3.22) и (3.23) l и d являются
средними величинами за сердечный цикл, поскольку до диаметра d=170 мкм имеет место
выраженная пульсация сосудов [189]; при d<170 мкм отношение диаметров в систолу и диастолу практически равно
1. Установлено [70], что при физической нагрузке прекапиллярные сосуды мышц расширяются незначительно или совсем не
расширяются. Следовательно, для терминальных микрососудов при изменении гипертензии
di(b) = di(bзс) , (3.24)
li(b) = li(bзс). (3.25)
Отметим, что несмотря на
гипертензию вязкость крови не изменяется как в “стандартных”, так и “нестандартных”
сосудах. Это происходит по следующим причинам. “Стандартные” сосуды “вписаны” в тот участок кривой, где
вязкость в сосудах постоянна и не зависит от диаметра (рис. 3.4). “Нестандартные”
сосуды приходятся на наклонный участок кривой, где величина вязкости крови зависит от диаметра сосудов, но
размеры последних при гипертензии практически не меняются. Следовательно, для
всякого i-сосуда при любой гипертензии
xi
(b) = xi(bзс). (3.26)
|
|
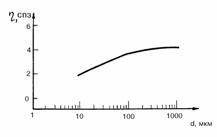
Рис. 3.4. Зависимость кажущейся вязкости от диаметра сосудов [185]. d -
диаметр сосуда, x - кажущаяся вязкость крови.
Очевидно, что для
“стандартных” сосудов при гипертензии
V i(b) = bV i(bзс), (3.27)
G i(b) = bG i(bзс). (3.28)
Кровоток в этих сосудах увеличивается в b2 раз [152]. Следовательно,
ti(b) = ti(bзс)/b, (3.29)
DPi(b) = bDPi(bзс).
(3.30)
На
“нестандартном” участке общий кровоток также возрастает в b2 раз. Однако здесь
существуют свои особенности. В “нестандартных” микрососудах цепочки и в капилляре кровоток при гипертензии увеличивается
только в b раз. Этот феномен обусловлен тем, что диаметр, длина и вязкость крови для
этих сосудов при гипертензии остаются неизменными. Установлено [251], что в покое в диапазоне d=19-50 мкм в
сердце перфузируется около 50% артериол; остальные составляют “резерв”. За счет
включения при гипертензии резервных микрососудов увеличивается плотность капиллярной сети сердца. Очевидно, что с
ростом физической нагрузки вниз по цепочке последовательно включаются резервные
сосуды все меньшего диаметра; тем самым увеличивается суммарная обменная поверхность “нестандартного” участка в
соответствии с уровнем гипертензии. За счет этого феномена также обеспечивается
плавное увеличение числа открытых капилляров, адекватное уровню нагрузки. Соотношение кровотоков в месте “подключения”
в постоянно открытом и резервном микрососудах следующее,
qi(b) = bqi (bзс), (3.31)
q*i
(b) = b(b-1)qi(bзс ), (3.32)
где qi(b), q*i (b) - соответственно кровотоки
в основном и резервном i-сосудах в заданном режиме гипертензии. Очевидно, что общий (основной+резервный)
кровоток возрастает в b2 раз. При этом в основном i-сосуде
Vi(b) = Vi(bзс ), (3.33)
Gi(b) = Gi(bзс ), (3.34)
DPi(b) = bDP i(bзс), (3.35)
t i(b) =
t i(bзс)/b, (3.36)
где Vi(b), Gi(b), DPi(b), t i(b) - объем, проводимость,
давление и время пребывания эритроцита в i-сосуде в данном режиме гипертензии. При
“задействовании” i-резервных сосудов общий объем и общая проводимость генераций
сосудов, расположенных ниже точки “подключения” и до капилляров, увеличивается
в b раз;
в b раз
возрастает и число открытых капилляров.
Таким образом,
с учетом вышеизложенного имеем два варианта закона (3.21):
1) для
“стандартных” сосудов
bDPi(bзс)t i (bзс )/b = 32x i(bзс )(b1/3(l i(bзс )/b1/3d i(bзс ))2,
2) для “нестандартных”
сосудов
bDPi (bзс)t i (bзс)/b = 32x i(bзс)(l i(bзс)/d i( зс))2
.
Каждое сечение терминальных
микрососудов и капилляров отдает кислород с постоянной скоростью как в покое,
так и при любом уровне гипертензии [152],
U(b) = 215%/с. (3.37)
Величина U(b)=215%/с - инвариант
микрососудистого и капиллярного участков сердца. Этот параметр не зависит ни от диаметра микрососудов, ни от уровня
гипертензии, ни от веса животных.
Поскольку при
гипертензии каждое сечение отдает кислород с постоянной скоростью, а время пребывания
эритроцитов в сосудах сокращается в b раз, то разность концентраций кислорода в крови на концах i-сосуда
DHbO2i (b) = DHbO2i (bзс )/b, (3.38)
где DHbO2i (b) - разность концентраций
кислорода в i-сосуде в заданном режиме гипертензии. Естественно, что разность концентраций на всем
терминальном участке также уменьшается в b раз; при этом напряжение кислорода на
артериальном конце капилляра возрастает на величину 42lgb [152],
apO2(b) = 50 + 42lgb.