3.3. Динамика зрительного образа
при аккомодации
Аккомодация есть автоматическое наведение оптики глаза на резкость изображения на сетчатке; автоматизм этого действия настолько ярко выражен, что, например, у человека, он даже практически неподвластен разуму и воле; сигналы, идущие от цилиарной мышцы, осуществляющей изменение округлости хрусталика глаза и тем самым — изменение его фокусного расстояния, не осознаются. Замечено, что аккомодация обнаруживается у животных (и у человека тоже) в самом раннем возрасте, и поэтому едва ли она в основе своей связана с процессом обучения. Рассмотрим динамику зрительного образа при наведении глаза на резкость и попытаемся объяснить таким образом механизм аккомодации.
Будем считать,
что при наведении глаза на резкость поток света, проходящий через его оптику,
сохраняется постоянным; и допустим, что сохраняется неизменной сумма
возбуждений фоторецепторов сетчатки: ![]() =const.
В действительности в силу нелинейностей рабочих характеристик рецепторов будет
некоторое несоответствие между изменением суммарного потока света и изменением
суммарного возбуждения рецепторов. Это несоответствие будет тем больше, чем
ближе к предельным окажутся сами возбуждения. Однако, если рассматривать
динамику зрительного образа вблизи исходного состояния, то погрешности будут
несущественными.
=const.
В действительности в силу нелинейностей рабочих характеристик рецепторов будет
некоторое несоответствие между изменением суммарного потока света и изменением
суммарного возбуждения рецепторов. Это несоответствие будет тем больше, чем
ближе к предельным окажутся сами возбуждения. Однако, если рассматривать
динамику зрительного образа вблизи исходного состояния, то погрешности будут
несущественными.
За основу
примем среднеарифметическое возбуждение рецепторов: ![]() , которое, согласно
принятым допущениям, сохраняется в процессе наведения на резкость постоянным;
и введём следующую зависимость преследующего возбуждения i-го рецептора
, которое, согласно
принятым допущениям, сохраняется в процессе наведения на резкость постоянным;
и введём следующую зависимость преследующего возбуждения i-го рецептора ![]() от предыдущего
(исходного)
от предыдущего
(исходного) ![]() :
:
![]() , (2.66)
, (2.66)
где k — коэффициент изменения резкости изображения на сетчатке глаза.
При k=1 возбуждения рецепторов сохраняются неизменными: ![]() =
=![]() ; при k=0 изображение на сетчатке глаза становится полностью
размытым, — и все возбуждения выравниваются:
; при k=0 изображение на сетчатке глаза становится полностью
размытым, — и все возбуждения выравниваются: ![]() =
=![]() ; при k>1 возбуждения возбуждённых выше
среднего рецепторов усилятся, а возбуждения возбуждённых ниже среднего — ещё
более ослабнут. Принятая зависимость (2.66)
не является единственной — возможны и
другие, — но мы остановимся на ней, так как она удовлетворяет нашим
требованиям.
; при k>1 возбуждения возбуждённых выше
среднего рецепторов усилятся, а возбуждения возбуждённых ниже среднего — ещё
более ослабнут. Принятая зависимость (2.66)
не является единственной — возможны и
другие, — но мы остановимся на ней, так как она удовлетворяет нашим
требованиям.
Наведение на
резкость будем оценивать по соотношению образов двух ситуаций: предыдущей и
последующей; при этом совершенно не определено, какой образ из этих двух более
резкий: первый или второю. Коэффициент приведения образа предыдущей ситуации ![]() к образу последующей
к образу последующей ![]() будет равен
будет равен
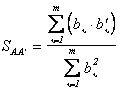 .
.
С учётом
зависимости (2.66) получим:
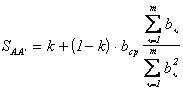 .
.Выражая
сумму возбуждений ![]() через среднеарифметическое
через среднеарифметическое ![]() , можно представить (2.67) в виде
, можно представить (2.67) в виде
![]() .
.
А если использовать сравнение образа исходной ситуации с единичным
образом, в частности — коэффициент
приведения образа к единичному: ![]() , — и встречный
коэффициент:
, — и встречный
коэффициент: ![]() =
=![]() , — то выражение (2.67) примет вид
, — то выражение (2.67) примет вид
![]() ,
,
или
![]() ,
,
где ![]() — степень сходства
образа А с единичным образом.
— степень сходства
образа А с единичным образом.
Встречный
коэффициент приведения последующего образа ![]() к предыдущему
к предыдущему ![]() с учетом (2.66)
определится как
с учетом (2.66)
определится как
![]() ,
(2.68)
,
(2.68)
или
![]() .
.
Коэффициент
приведения любого из рассматриваемых образов — предыдущего или последующего — к
самому себе через
другой, т.е. степень сходства этих образов, равен произведению коэффициентов ![]() и
и ![]() :
:
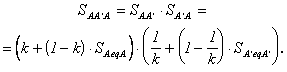 (2.69)
(2.69)
|
Рис.2.6. Динамика зрительного образа при наведении глаза на резкость |
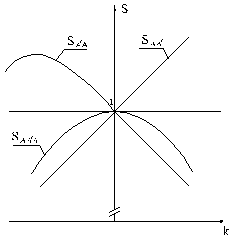
Полученные
зависимости (2.67), (2.68) и (2.69)
отображены на рис. 2.6. Коэффициент приведения
образа предыдущей ситуации к образу последующей ![]() имеет линейную
зависимость от изменения резкости изображения, характеризуемого коэффициентом k, причём при возрастании резкости изображения (k>1)
указанный коэффициент приведения становится больше единицы, а при снижении резкости изображения (k<1) — меньше единицы; такая особенность может быть положена в основу
объяснения аккомодации, но об этом — чуть позже.
имеет линейную
зависимость от изменения резкости изображения, характеризуемого коэффициентом k, причём при возрастании резкости изображения (k>1)
указанный коэффициент приведения становится больше единицы, а при снижении резкости изображения (k<1) — меньше единицы; такая особенность может быть положена в основу
объяснения аккомодации, но об этом — чуть позже.
Зависимость
встречного коэффициента ![]() от изменения резкости
изображения — более сложная. В некотором
смысле этот коэффициент как бы обратный по отношению к первому: при возрастании
резкости (k>1) он — меньше единицы, а при снижении (k<1) — больше. Но при полностью размытом (нерезком)
изображении на сетчатке глаза в последующей ситуации, когда k стремится к нулю, коэффициент приведения
от изменения резкости
изображения — более сложная. В некотором
смысле этот коэффициент как бы обратный по отношению к первому: при возрастании
резкости (k>1) он — меньше единицы, а при снижении (k<1) — больше. Но при полностью размытом (нерезком)
изображении на сетчатке глаза в последующей ситуации, когда k стремится к нулю, коэффициент приведения ![]() приближается к
единице, т.е. размытый образ как будто не имеет лица и в приведении не
участвует
приближается к
единице, т.е. размытый образ как будто не имеет лица и в приведении не
участвует
Кривая степени
сходства образов предыдущей и последующей ситуаций ![]() располагается на
графике (рис.2.6) всегда ниже уровня
единицы, и, чем дальше расходятся образы по изменению резкости изображения, тем
меньше становится эта степень, однако до нуля она не доходит: некоторое
сходство ситуаций (точнее — их образов)
всё же сохраняется. Если образ последующей ситуации полностью размыт (k=0) , то степень сходства становится равной
коэффициенту приведения
располагается на
графике (рис.2.6) всегда ниже уровня
единицы, и, чем дальше расходятся образы по изменению резкости изображения, тем
меньше становится эта степень, однако до нуля она не доходит: некоторое
сходство ситуаций (точнее — их образов)
всё же сохраняется. Если образ последующей ситуации полностью размыт (k=0) , то степень сходства становится равной
коэффициенту приведения ![]() . Можно ещё отметить то, что в зоне большей резкости (k>1) кривая степени
сходства более пологая; это означает, что при увеличении резкости изображения
образ изменяется меньше, чем при снижении её.
. Можно ещё отметить то, что в зоне большей резкости (k>1) кривая степени
сходства более пологая; это означает, что при увеличении резкости изображения
образ изменяется меньше, чем при снижении её.
А теперь вернёмся к объяснению аккомодации; это объяснение необходимо нам для того, чтобы можно было найти техническое решение автоматического наведения на резкость технического глаза обучаемой системы управления. Можно, разумеется, обойтись без объяснений, если подходить к задаче создания механизма аккомодации логическим путём, впрочем, такие механизмы уже созданы, и применяются они для автоматического наведения на резкость фотоаппаратов. В основе их работы, как правило, лежит использование нелинейностей рабочих характеристик фотодатчиков. Изображение проецируется на экран, состоящий из множества фотодатчиков. При наведении изображения на резкость освещение одних датчиков усиливается, а других — ослабевает; сигналы первых увеличиваются, а других уменьшаются. Принимая во внимание, что рабочие характеристики датчиков нелинейные, а точнее: при усилении освещения сигналы на их выходах изменяются логарифмически (или экспоненциально), т.е. стремятся к своему пределу, то увеличение суммарного — сигнала первых фотодатчиков, тех, освещение которых усиливается, будет не столь большим, как уменьшение других; в результате суммарный сигнал фоточувствительного экрана при наведении на резкость будет уменьшаться, а при размывании изображения, наоборот, — увеличиваться. Далее известными средствами автоматики решается задача минимизации сигнала управления.
Попытаемся предложить иной принцип работы механизма аккомодации, опираясь на динамику образов при наведении на резкость. Но прежде отметим, что едва ли аккомодации можно обучить: просто нет таких ситуаций, которые характеризовали бы резкость изображения; а коли нет характерных ситуаций, нет и никаких надежд на обучение. Впрочем, классическая нейрофизиология однозначно считает аккомодацию врождённым рефлексом [17]. Но ведь и у врождённого рефлекса должен быть свой принцип действия: каков он? на чём он основывается? Можно даже уточнять вопрос: каким образом рефлекс, т.е. система управления, оценивает резкость изображения на сетчатке глаза?
Ответ будем
искать, как условились, в динамике образов. Прежде всего отметим, что при любом
изменении резкости образы возбуждений рецепторов будут отличаться друг от
друга, о чём свидетельствует степень сходства ![]() (рис.2.6).
Этого уже достаточно для того, чтобы утверждать, что, если есть отличие образов,
то можно это отличие использовать для управления механизмом аккомодации.
(рис.2.6).
Этого уже достаточно для того, чтобы утверждать, что, если есть отличие образов,
то можно это отличие использовать для управления механизмом аккомодации.
Более
конкретно о принципе управления можно говорить, основываясь на зависимости
коэффициента приведения предыдущего образа к последующему ![]() от изменения резкости
изображения k. То, что этот коэффициент больше
единицы при усилении резкости и меньше —
при снижении её ( рис.2.6),
можно воспринять как ключ к разгадке секрета аккомодации. Осталось только дополнить
наши рассуждения тем, что указанный коэффициент приведения определяет
соотношение фактических сигналов управления в двух следующих одна за другою
ситуациях при условии, что проводимости синапсов формируются в предыдущей из
них; об этом говорит выражение (2.14). Стыкуя то и другое, получим, что при
росте резкости (k>1)
коэффициент приведения
от изменения резкости
изображения k. То, что этот коэффициент больше
единицы при усилении резкости и меньше —
при снижении её ( рис.2.6),
можно воспринять как ключ к разгадке секрета аккомодации. Осталось только дополнить
наши рассуждения тем, что указанный коэффициент приведения определяет
соотношение фактических сигналов управления в двух следующих одна за другою
ситуациях при условии, что проводимости синапсов формируются в предыдущей из
них; об этом говорит выражение (2.14). Стыкуя то и другое, получим, что при
росте резкости (k>1)
коэффициент приведения ![]() будет больше единицы
(
будет больше единицы
(![]() >1), и, если реализовать зависимость (2.14), то фактический
сигнал управления в последующей ситуации
>1), и, если реализовать зависимость (2.14), то фактический
сигнал управления в последующей ситуации ![]() окажется больше сигнала
окажется больше сигнала
![]() в предыдущей:
в предыдущей: ![]() и
и ![]() >
>![]() . И наоборот, при k<1 получим
. И наоборот, при k<1 получим ![]() >1
и
>1
и ![]() >
>![]() . Другими словами: при усилении резкости изображения
сигнал управления будет увеличиваться, а при снижении — уменьшаться.
. Другими словами: при усилении резкости изображения
сигнал управления будет увеличиваться, а при снижении — уменьшаться.
Напомним, что
указанная связь между сигналами управления и образами возникает при выполнения
двух важнейших условий: во-первых, необходимо постоянное изменение образа,
т.е. необходимо постоянное движение механизма
наведения на резкость; стоит только остановиться ему, и та динамика образа,
которая лежит в основе принципа действия, потеряет
свою силу; во-вторых, мы должны признать, что аккомодация как рефлекс всё же нуждается в обучении, но не в обычном обучении,
а как бы в скользящем, когда оно всякий раз
осуществляется в предыдущей ситуации; только при этом условии оказывается
справедливой зависимость, отражённая в выражении
(2.14).
Приняв оба эти
условия, мы должны подумать о том, как они реализуются на самом деле.
Непрерывное движение механизма наведения на резкость легко обеспечивается
нестабильностью системы регулирования. Из теории автоматического управления известно,
что любая система с отрицательной обратной связью склонна к колебаниям по своей
природе, тем более, если этому способствуют её параметры, в частности такой,
как запаздывание обратной связи. В нашем случае датчиками обратной связи являются
рецептор сетчатки, а они, как известно, срабатывают, т.е. выдают на выходе
сигнал, не мгновенно; способствовать неустойчивости могут и другие параметры
системы, например инерционность. Так или иначе, но неустойчивость, о которой мы
ведём речь, не только может быть, но и есть в действительности, о чём свидетельствуют
исследования в области офтальмологии [21].
Что же касается скользящего обучения системы управления, то объяснить его можно только с учётом особого режима, в котором есть забывание, но нет запоминания. Вспомним в связи с этим ещё раз наше толкование процесса обучения мозга животных; этот процесс состоит из трёх элементов: 1) увеличение проходных сечений синапсов под воздействием проходящих через них своих управляющих потоков — так называемое саморегулирование; 2) запоминание, т.е. стимуляция роста проводимостей синапсов и фиксация этих проводимостей с помощью особых медиаторов-стимуляторов запоминания и, наконец, 3) забывание, т.е. зарастание проходных сечений синапсов под воздействием других медиаторов-стимуляторов забывания. Если исключить из процесса обучения второй элемент, то есть подкрепление увеличения проходных сечений синапсов путём воздействия медиаторов запоминания, то память окажется кратковременной, определяемой своего рода упругой деформацией синапсов.
Ранее мы
договорились считать, что стимуляторы запоминания выделяются в нейронах при
положительных эмоциях, т.е. при одобрении определённых действий, но, как
известно из нейробиологии, действия цилиарной мышцы человеком никак не ощущаются
и поэтому никак не оцениваются; а это значит, что одобрить её действия выделением
стимуляторов запоминания человек бессилен. Остаётся согласиться с тем, что
синапсы управления наведением на резкость, увеличивающие свои проводимости под
воздействием протекающих через них управляющих потоков, способны сохранить эти
проводимости на очень короткое время, определяемое последействием синапсов
или, что одно и то же, скоростью забывания. Конечно, время полного забывания у
всех синапсов различное, но его можно осреднить и считать одинаковым; пусть это
время будет равно ![]() : именно это время
: именно это время ![]() разделяет предыдущую и
последующую ситуации.
разделяет предыдущую и
последующую ситуации.
На рис. 2.7 представлена схема одной из возможных
систем управления аккомодацией о явлением постоянного забывания. Объяснение
можно начать с любой текущей ситуации, в которой возбуждение i-го рецептора равно ![]() . Это возбуждение формирует проводимость соответствующего
синапса:
. Это возбуждение формирует проводимость соответствующего
синапса: ![]() , где k — коэффициент
пропорциональности обучения. Спустя время
, где k — коэффициент
пропорциональности обучения. Спустя время ![]() возбуждение того же
рецептора изменится и станет равным
возбуждение того же
рецептора изменится и станет равным ![]() , но проводимость
соответствующего синапса в силу своего последействия сохранится предыдущим, то
есть равным
, но проводимость
соответствующего синапса в силу своего последействия сохранится предыдущим, то
есть равным ![]() .
.
Рецепторная
доля сигнала управления ![]() , определяемая возбуждением
, определяемая возбуждением ![]() и прежней
проводимостью
и прежней
проводимостью ![]() , будет равна:
, будет равна: ![]() . Постепенно возбуждения
. Постепенно возбуждения ![]() изменят проводимость
синапса и сделают её равной
изменят проводимость
синапса и сделают её равной ![]() , так что, спустя
очередной отрезок времени
, так что, спустя
очередной отрезок времени ![]() возбуждение
рассматриваемого рецептора уже станет равным
возбуждение
рассматриваемого рецептора уже станет равным ![]() , а его доля
—
, а его доля
— ![]() , и так далее. Если теперь просуммировать
предыдущие доли
, и так далее. Если теперь просуммировать
предыдущие доли ![]() :
:
![]() ,
,
- а затем просуммировать те же доли в последующей ситуации:
![]() ,
,
то, вычитая
одну сумму из другой, получим разность ![]() как текущий сигнал управления механизма
наведения на резкость. Эта разность будет положительной при наведении на
резкость и отрицательной при уходе с резкости, при размывании изображения на
сетчатке.
как текущий сигнал управления механизма
наведения на резкость. Эта разность будет положительной при наведении на
резкость и отрицательной при уходе с резкости, при размывании изображения на
сетчатке.
После объяснения принципов построения системы регулирования механизма наведения на резкость рассмотрим его работу. Цилиарная мышца представляет собой привод одностороннего действия; она делает хрусталик глаза более выпуклым. Хрусталик ведёт себя при этом как пружина; обратной мышцы или как её ещё называют — антагонистической у цилиарной нет. В исходном состоянии, когда нет сигнала управления и мышца не действует, хрусталик полностью распущен, уплощён, то есть наведён на резкость в бесконечности; такую форму он приобретает сам в силу своей упругости. После включения в работу на цилиарную мышцу подаётся сигнал управления и она начинает сокращаться, делая хрусталик более выпуклым. Пока будет обостряться резкость изображения сигнал управления, согласно нашим выводам, будет больше нуля и мышца будет продолжать сокращаться. При выходе на наивысшую резкость сигнал должен был бы уменьшиться до нуля, но этого не произойдёт по той простой причине, что и рецепторы, и синапсы действуют с запаздыванием. Сигнал управления уменьшится до нуля спустя время запаздывания; и только тогда цилиарная мышца отключится и расслабится. Упруго напряжённый хрусталик начнёт деформироваться в обратную сторону, уменьшая свою кривизну. Допустим, что отключение цилиарной мышцы продолжается всегда на одно и то же время, достаточное для того, чтобы хрусталик перескочил в обратную сторону через точку наивысшей резкости; и когда она снова подключится, её сигнал окажется положительным, снова она начнёт сжимать хрусталик, и снова изображение начнёт увеличивать свою резкость. Движение повторится и будет повторяться постоянно раз за разом пока глаз смотрит на объект. Таков в общих чертах механизм аккомодации глаза, точное говоря — один из возможных вариантов механизма; и по такому принципу может действовать реальный механизм наведения на резкость, встроенный в обучаемую систему управления.
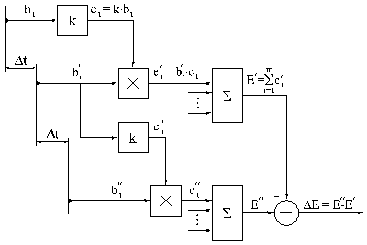
Рис.2.7. Схема автоматической системы управления механизма наведения на резкость
Элемент сравнения, на котором происходит вычитание предыдущего сигнала управления из последующего (рис.2.7), может отсутствовать при условии, что есть утечка сигнала управления; при этом условии сигнал управления появляется тогда, когда сигнал на выходе из сумматора будет иметь величину, большую чем утечки.
В заключение приведём параметры аккомодации человеческого глаза; если расстояние до объекта соответствует 1,0 диоптрию, а диаметр зрачка равен 7 мм, то колебания совершаются с частотой 1... 3 Гц и амплитудой 0,2...0,3 диоптрия. При уменьшении диаметра зрачка, то есть при увеличении глубины резкости, и при удалении объекта наблюдения колебания уменьшают свою амплитуду.